Blastomeri: prime fasi dello sviluppo embrionale
Subito dopo la fecondazione, lo zigote umano inizia a dividersi in blastomeri, cellule piccole ma decisive per il destino dell’embrione. In queste ore iniziali si concentra un’enorme quantità di informazioni biologiche, che orienta l’intero sviluppo successivo.
Lo zigote deriva dall’unione di ovocita e spermatozoo, entrambi prodotti tramite meiosi.
Il suo genoma rimane ancora silente e le prime divisioni, dette clivaggio, utilizzano soprattutto riserve materne. Le cellule che si formano non aumentano il volume complessivo dell’embrione, ma ne moltiplicano il numero cellulare.
In questo modo si impostano le basi per la futura organizzazione di tessuti e organi. Comprendere come agiscono questi primi blastomeri è cruciale per la biologia dello sviluppo e per la medicina della riproduzione assistita, che si confronta quotidianamente con tali passaggi.
Il tema ha anche un forte risvolto clinico. Alterazioni nelle prime divisioni possono compromettere impianto e prosecuzione della gravidanza, oppure generare instabilità cromosomica.
Chi studia embriologia, genetica o tecniche di laboratorio incontra spesso il termine e deve collegarlo a eventi molecolari specifici.
Nei punti successivi analizzeremo origine, cronologia delle divisioni, compattazione e formazione della blastocisti. Vedremo poi le funzioni chiave, la relazione con le cellule staminali e il significato clinico di asimmetrie e anomalie, collegando i dati sperimentali alle applicazioni pratiche.
Definizione cellulare e origini dei blastomeri
I blastomeri sono le prime cellule che compaiono dopo la formazione dello zigote e rappresentano l’avvio concreto dello sviluppo embrionale. Ogni cellula deriva da una divisione mitotica rapida, priva della classica fase di crescita intermedia che caratterizza altri tipi di cicli cellulari.
Dopo la fecondazione, il nucleo dello zigote contiene un corredo cromosomico diploide, ottenuto grazie alla precedente meiosi nei gameti parentali.
Le prime divisioni di clivaggio sono inizialmente sincrone e producono 2, 4, poi 8 cellule. In questa fase le cellule restano totipotenti, cioè capaci di dare origine a un organismo intero, con tessuti embrionali ed extraembrionali.
La divisione richiede un controllo accurato del fuso mitotico e del cinetocore, struttura essenziale per la corretta segregazione dei cromosomi.
Qualsiasi errore in questi meccanismi può avere conseguenze permanenti, perché interviene quando il numero di cellule è ancora molto limitato.
Per visualizzare il ruolo dei blastomeri, si può pensare a una catena di montaggio: ogni divisione moltiplica i “moduli” senza cambiare le dimensioni complessive del prodotto. Il processo ricorda la mitosi, ma avviene in un contesto estremamente regolato e con cicli abbreviati.
Errori precoci possono generare mosaicismi cromosomici già agli stadi iniziali. Queste cellule costituiscono quindi il punto di partenza obbligato per la formazione dei foglietti germinativi, da cui deriveranno in seguito tessuto epiteliale, tessuto connettivo e le altre grandi categorie tissutali dell’organismo adulto.
Divisioni dei blastomeri: dallo zigote alla morula
La dinamica temporale delle divisioni dei blastomeri è sorprendentemente precisa nell’uomo. La prima divisione dello zigote avviene, in media, tra 24 e 30 ore dopo la fecondazione, segnando l’ingresso nello stadio a due cellule.
Da questo momento seguono altre divisioni di clivaggio, inizialmente ancora abbastanza sincrone. Entro il terzo giorno post-fecondazione, l’embrione raggiunge lo stadio di morula, formato da circa 8–16 blastomeri.
In questa fase il volume complessivo rimane costante, mentre il diametro delle singole cellule diminuisce progressivamente ad ogni ciclo mitotico.
Le cellule appaiono sempre più strettamente impacchettate, anche se la compattazione vera e propria avverrà solo poco dopo. È importante ricordare che, nei mammiferi, le divisioni diventano progressivamente asincrone, quindi il numero di cellule non segue più in modo rigoroso potenze di due.
Immagina di osservare, con un sistema time-lapse, una sequenza temporale delle prime ore: si partirebbe da due cellule quasi identiche, per arrivare rapidamente a un ammasso compatto. In poche decine di ore cambia completamente la geometria dell’insieme.
Questa cronologia è cruciale, per esempio, nella valutazione degli embrioni in fecondazione in vitro.
Un embrione che non raggiunge correttamente lo stadio di morula entro i tempi attesi mostra spesso un potenziale di sviluppo ridotto. Lo studio sistematico delle tempistiche di divisione permette di collegare dati morfologici e probabile esito dell’impianto.
Compattazione, blastocisti e prime linee cellulari distinte
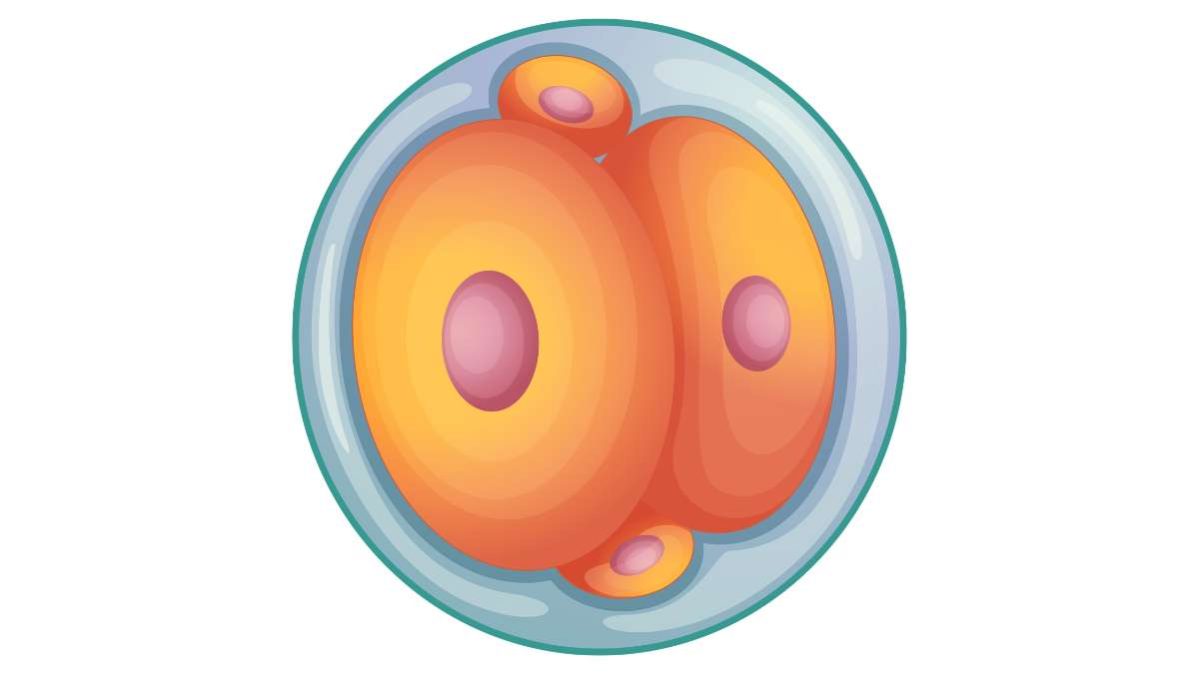
Raggiunti gli 8–16 blastomeri, l’embrione entra nella fase di compattazione.
Le cellule superficiali si appiattiscono, stabiliscono giunzioni strette e formano un involucro continuo che racchiude una massa interna meno esposta.
Tra il terzo e il quarto giorno, la morula entra nella cavità uterina.
Le cellule pompano attivamente ioni mediante pompe sodio‑potassio, richiamando acqua negli spazi intercellulari.
Si forma così una cavità piena di fluido, il blastocele, che separa il gruppo interno di cellule dall’involucro più esterno.
Tra il quinto e il sesto giorno compare la blastocisti, con diametro di circa 0,1–0,2 mm e un numero complessivo di 100–200 cellule dopo 7–8 divisioni.
A questo punto si individuano due popolazioni: la inner cell mass, destinata all’embrione vero e proprio, e il trophectoderm, che formerà placenta e membrane extraembrionali.
I blastomeri non sono più equivalenti; iniziano le prime decisioni di destino cellulare. Differenze di posizione, polarità e contatti di membrana guidano l’attivazione differenziale dei geni, trasformando piccole disuguaglianze in traiettorie di sviluppo divergenti.
Queste scelte precoci avranno ripercussioni profonde, perché determineranno la formazione successiva dei foglietti germinativi e quindi di tessuto epiteliale, tessuto connettivo e altri tessuti specializzati.
In questa finestra temporale, errori strutturali o funzionali possono impedire il corretto impianto della blastocisti nell’endometrio, riducendo drasticamente le possibilità di una gravidanza evolutiva.
Totipotenza, genoma embrionale e relazione con le cellule staminali
Nelle prime divisioni, i blastomeri sono definiti totipotenti.
Ogni cellula possiede cioè la capacità teorica di generare un organismo completo, inclusi i tessuti extraembrionali che sosterranno l’embrione durante la gestazione.
Questa totipotenza si mantiene almeno fino allo stadio di morula.
Dal punto di vista funzionale, le cellule passano da una fase dominata da mRNA materni a una fase in cui si attiva il genoma embrionale. Questo processo, noto come embryonic genome activation, avviene tipicamente durante la transizione da 4 a 8 cellule.
In quel momento cambia profondamente il profilo di espressione genica e la regolazione della sintesi proteica. Le proprietà dei blastomeri richiamano alcuni aspetti delle cellule staminali, pur collocandosi in una fase ancora più primitiva e con un ventaglio di possibilità maggiore rispetto alla pluripotenza classica.
Ecco i principali attributi funzionali dei blastomeri precoci:
- Elevata totipotenza e plasticità di destino cellulare
- Rapido ciclo cellulare con divisioni di tipo mitotico
- Forte dipendenza iniziale da trascritti materni preformati
- Progressiva autonomia grazie all’attivazione del genoma embrionale
Per chi studia cellule staminali, questi stadi rappresentano un modello estremo di plasticità biologica.
Comprendere come la totipotenza si restringe progressivamente aiuta a interpretare i passaggi che portano alla pluripotenza e poi alla specializzazione finale.
Inoltre, il controllo fine della sintesi proteica in questa fase offre un riferimento per molte strategie di medicina rigenerativa.
Asimmetrie di divisione, instabilità cromosomica e impatto clinico
Non tutti i blastomeri si dividono in modo perfettamente simmetrico.
Nei mammiferi, le divisioni diventano progressivamente asincrone, con numeri cellulari irregolari e talvolta con dimensioni molto eterogenee tra una cellula e l’altra.
Si parla di unequal cleavage quando una divisione produce cellule molto diverse per volume. Studi clinici hanno mostrato che queste divisioni non simmetriche riducono il tasso di impianto, che scende intorno al 23,9–36,4%, e il tasso di gravidanza, stimato tra 37,6–52,9%, rispetto agli embrioni con divisioni regolari.
In condizioni ancora più estreme può verificarsi reverse cleavage, cioè fusione di blastomeri o mancata citodieresi dopo una divisione nucleare. In uno studio, solo circa il 9% degli embrioni con reverse cleavage ha raggiunto la blastocisti, indicando una compromissione severa del potenziale di sviluppo.
A livello molecolare, queste anomalie riflettono spesso instabilità cromosomica. Errori di segregazione durante la mitosi, legati anche a difetti del cinetocore, producono cellule aneuploidi o mosaicismi complessi, difficili da correggere in fasi successive.
Analisi di embrioni umani in fecondazione in vitro hanno evidenziato riarrangiamenti cromosomici già negli stadi iniziali di clivaggio.
I blastomeri diventano così un indicatore precoce dell’affidabilità genetica dell’embrione. Interpretare correttamente pattern di divisione, dimensioni cellulari e tempi di clivaggio è essenziale per collegare morfologia, genotipo e reale potenziale di sviluppo.
Significato dei blastomeri nella visione dello sviluppo umano
Osservare i blastomeri significa assistere alla prima organizzazione visibile del progetto umano.
In poche divisioni controllate si passa da una singola cellula a una struttura complessa, capace di differenziarsi in tutti i tessuti.
Dalla definizione iniziale, attraverso la cronologia di clivaggio, la compattazione e la formazione della blastocisti, emerge un quadro coerente. Ogni passaggio richiede coordinazione tra cicli cellulari, stabilità cromosomica e regolazione fine della sintesi proteica.
È in questo crocevia che si decidono il successo dell’impianto e la corretta formazione di tessuto epiteliale, tessuto connettivo e di ogni altro distretto corporeo.
La totipotenza dei blastomeri, pur effimera, rappresenta il vertice massimo della plasticità cellulare, più ampia persino di quella delle classiche cellule staminali.
Questa architettura biologica non assomiglia alla bioarchitettura umana solo per analogia: entrambe mostrano come ordine e complessità possano emergere da regole relativamente semplici, applicate con rigore.
Studiare i blastomeri offre quindi una lente privilegiata sulla logica profonda che guida lo sviluppo, dalla prima divisione dello zigote fino alla complessità dell’organismo adulto.